Archives
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2018-07
-
The swelling independent activation of
2021-08-13
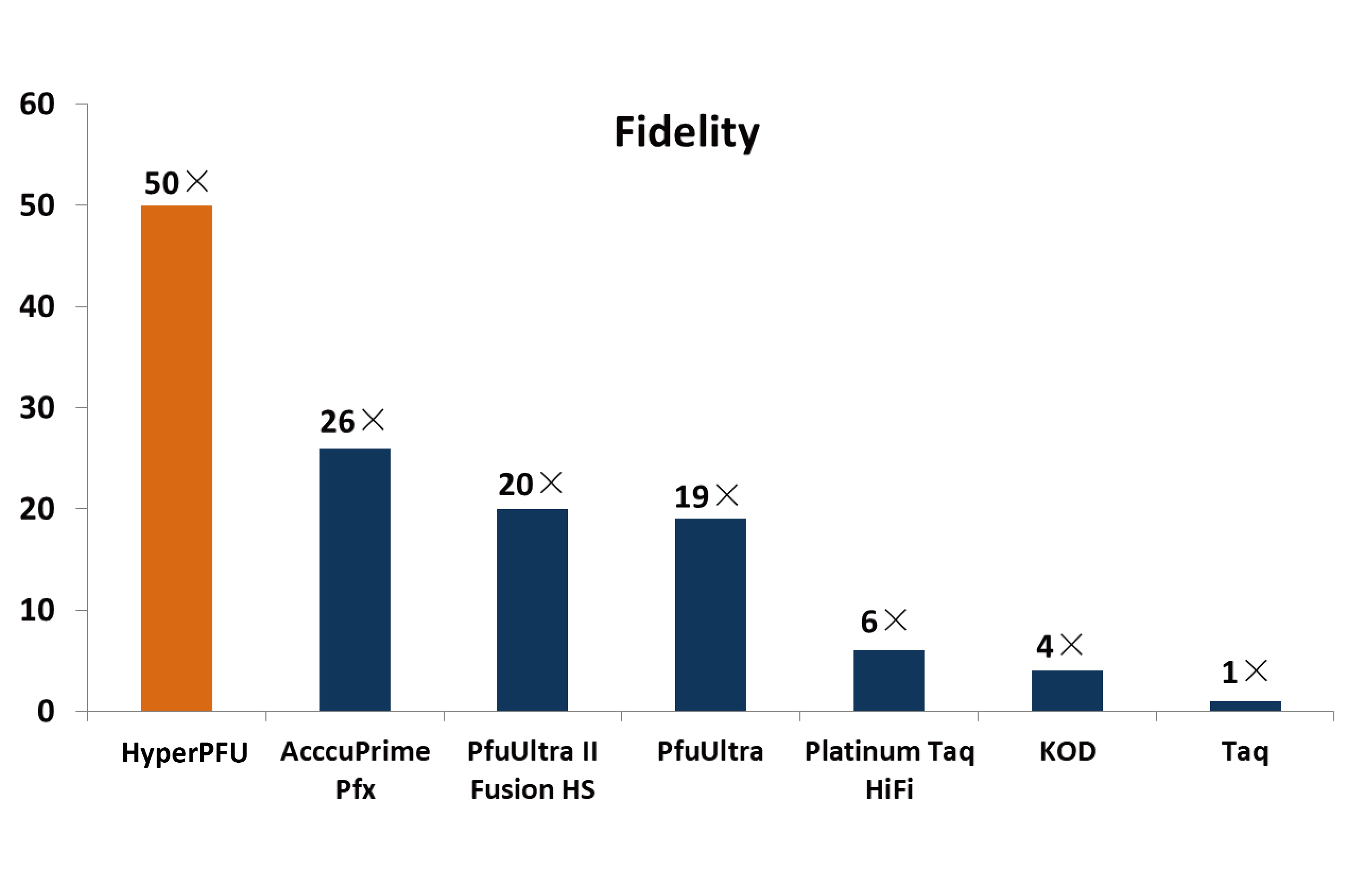
The swelling-independent activation of chloride channels by glycerol is proved by our further experiments. When glycerol concentration in the isoosmotic solution was reduced to the micromolar scale or even lower concentration, a Cl− current was still induced. The Cinobufagin were not swollen by the
-
Plant uptake of P from OA could
2021-08-13
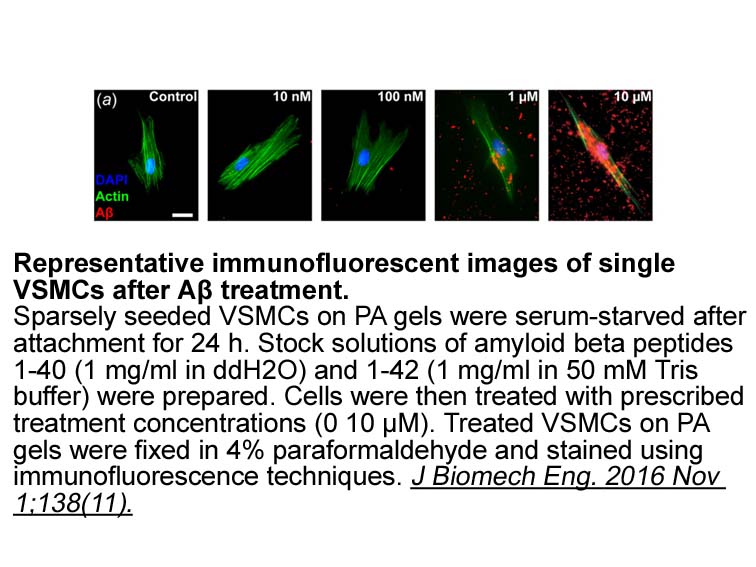
Plant uptake of P from OA could also be affected by arbuscular mycorrhizal fungi (AMF). Approximately 80% of terrestrial plant species form symbiotic associations with AMF (Smith and Read, 2010) which can enhance plant P uptake (Facelli and Facelli, 2002, Pearson and Jakobsen, 1993, Tibbett, 2000).
-
So what can we glean from this illuminative foray into
2021-08-13
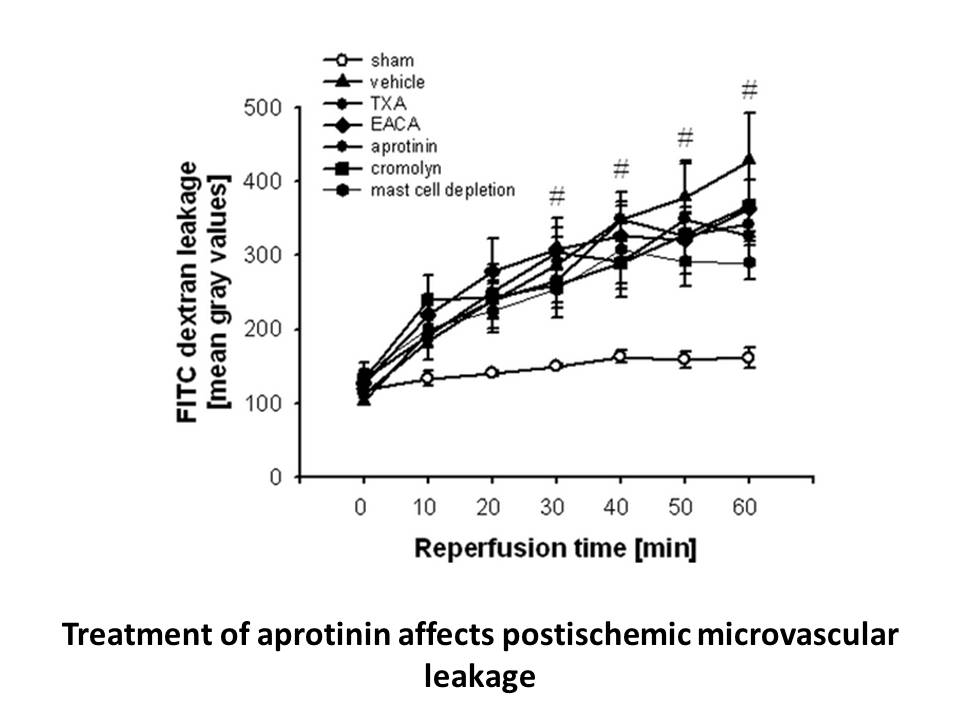
So what can we glean from this illuminative foray into how Chk1 and MK2 participate in checkpoint control? First and foremost, this work reveals that microcystin lr deficient in the tumor suppressor p53 contain two spatially distinct G2/M phase checkpoint control kinase networks. This provides comp
-
br O GlcNAc transferase OGT belongs to the metal independent
2021-08-12

O-GlcNAc transferase OGT belongs to the metal-independent GT-B superfamily of glycosyltransferases, which has been well-reviewed previously [14,15]. OGT is an essential gene encoded on the X-chromosome, and it has two main regions: a long N-terminal tetratricopeptide repeat (TPR) region and a C-t
-
Enzyme selectivity of many enzymes like galactosidases acyla
2021-08-12
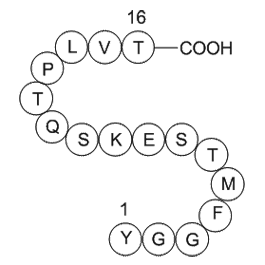
Enzyme selectivity of many purchase roscovitine like β-galactosidases, acylases, epoxyhydrolases, lipases, etc. has also been modulated by using different immobilization techniques. It can result in the selection of optimal biocatalyst for different processes. This has many advantages over strategi
-
br Acknowledgements br Introduction Supramolecular collagen
2021-08-12

Acknowledgements Introduction Supramolecular collagen assemblies are crucial for the mechanical stability of animal bodies (Myllyharju and Kivirikko, 2004). The basic collagen structure is a triple helix of three chains containing multiple Gly-X-X' repeats; X and X' are often proline and 4-hyd
-
Herbal products may alter the metabolism of
2021-08-12
Herbal products may alter the metabolism of certain drugs and, likewise, the metabolism of components of herbal products may be altered by some drugs, specifically those that affect cytochrome P450 metforman (CYP). These can bring about an alteration of the pharmacokinetics and pharmacological acti
-
GANT61 Contrary to the downregulation of S PR
2021-08-12
Contrary to the downregulation of S1PR family GPCRs, CLL cells have increased mRNA expression of the lysophosphatidic GANT61 (LPA) family receptors LPAR1, LPAR3 and LPAR4 compared to normal B cells [47]. Increased LPAR1 mRNA has been shown to be associated with more aggressive disease [47] and LPA
-
It was hypothesized that if modafinil acts primarily via
2021-08-12

It was hypothesized that if modafinil acts primarily via a noradrenergic mechanism, Dbh −/− mice should be non-responsive since they completely lack NE. In contrast, if modafinil acts mainly through DA systems, these mice should be hypersensitive. Modafinil was tested in Dbh −/− mice using both loco
-
In the present work we have investigated the reactivity of
2021-08-12
In the present work, we have investigated the reactivity of N-aryl-N′-hydroxyguanidines 1a–d (see Scheme 1 for structures) with the water-soluble Cu(II) complex 8. Using EPR and UV–Visible spectroscopy, we have shown that the studied N-aryl-N′-hydroxyguanidines 1a–d can bind and transfer electrons t
-
br Methods br Results br
2021-08-12
Methods Results Discussion To our knowledge, this is the first report to investigate the association between DBH genotype and pDβH activity in Han Chinese subjects. We also examined the relationship among DBH gene polymorphisms, pDβH activity and cognitive function in patients with schizoph
-
DNA methylation is associated with transcriptional silencing
2021-08-12
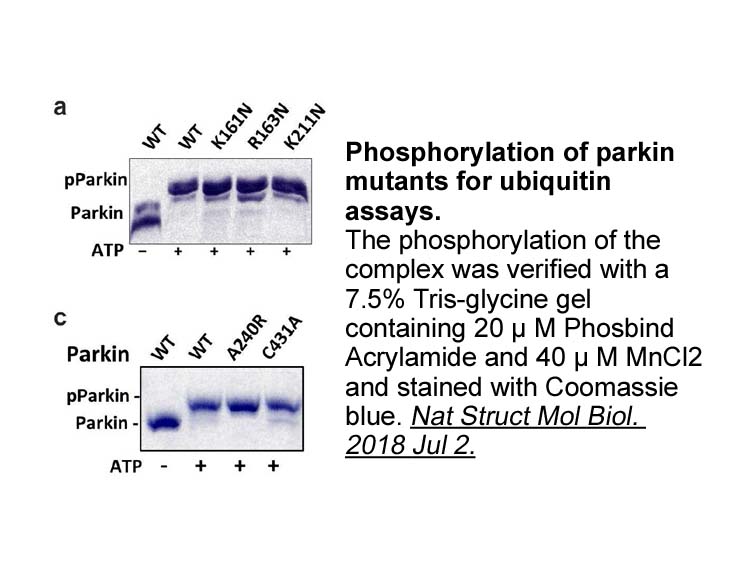
DNA methylation is associated with transcriptional silencing of tumor suppressors or other genes important for normal cellular function and plays an important role in the development of cancer and other diseases (such as breast and colorectal cancers) [19]. DNA methyltransferases (MTases) are a fami
-
In our recent study we established a procedure to examine
2021-08-12
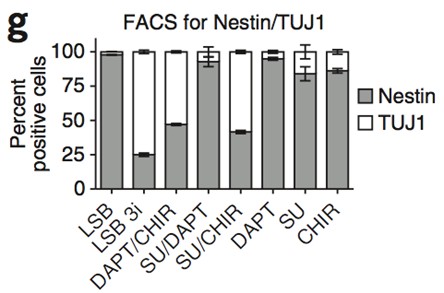
In our recent study, we established a procedure to examine the chromatin binding of XRCC4 using a biochemical fractionation analysis using a detergent Nonidet P-40 [22]. In this study, we investigated the role of LIG4 and its subdomains in the recruitment of XRCC4/LIG4 complex to chromatin. Mater
-
The possibility that a carboxyl group attached to the
2021-08-12
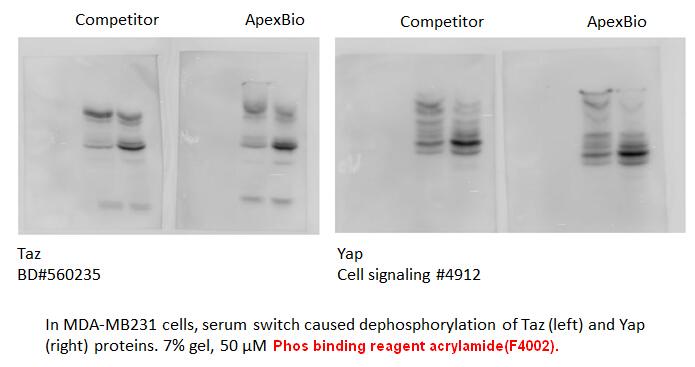
The possibility that a carboxyl group attached to the or position of an -arylaminomethylenemalonate ester would lead to increased selectivity for DHODH has been tested and found not to be the case. Indeed, reduced selectivity compared with the corresponding ester is observed and is attributed to a ‘
-
zotarolimus br Materials and methods br Results br Discussio
2021-08-12
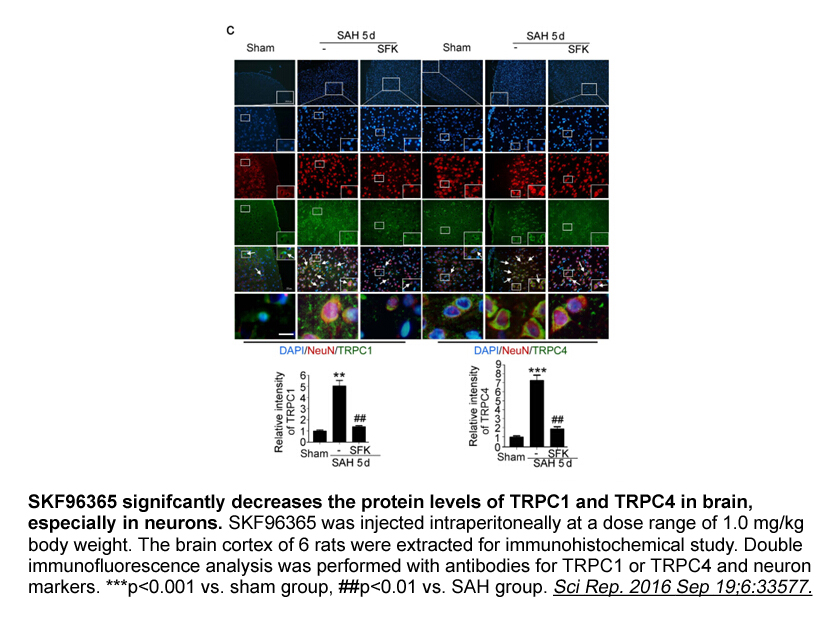
Materials and methods Results Discussion Cysteine proteases are upregulated under oxidative stress and play an important role in the preservation of cellular metabolism (Usui et al., 2007). Cysteine proteases are also important under abiotic stresses for plants to degrade proteins denatured
14902 records 620/994 page Previous Next First page 上5页 616617618619620 下5页 Last page